In December 2019, a series of acute respiratory illnesses characterized by rapid worsening of symptoms, pneumonia, acute respiratory distress syndrome (ARDS), and in some cases, death were observed in Wuhan City, Hubei Province, China. To achieve the set objective, the results of treatment of 100 sexually mature rats of both sexes with coronavirus infection caused by COVID -19 were analyzed. All animals were divided into groups: Group 1 - animals with coronavirus infection with a confirmed positive PCR test, treated with ivermectin at a dosage of 300 mg of body weight (n = 25), Group 2 - animals with coronavirus infection treated with baicalin at a dosage of 500 mg (n = 25), Group 3 - animals with coronavirus infection treated with molnupiravir 25 mg / kg of body weight (n = 25), Group 4 - animals with coronavirus infection treated with a new drug based on G. lucidum and black cumin (n = 25). Lymphocytic myocarditis was detected in all rats (100%). Dystrophic changes in cardiomyocytes were also detected in 57.4% of cases, apoptotic bodies in 29.41% of cases, uneven cardiomyocyte hypertrophy in 85.29% of cases, and lipofuscinosis in 45.59% of cases. Subendocardial lipomatosis in the cardiac conduction system was observed in 51.47% of cases. The average percentage of lipomatosis area from the micrograph area was 2.47±3.12%. The area under the curve (AUC) = 0.91 (95% CI = 0.85-0.98). The prediction accuracy of the prognostic model was 91.7%, sensitivity was 0.87%, and specificity was 0.95%. AUC was 0.91 (95% CI = 0.85-0.98), p < 0.001.
| Published in | American Journal of Medical Science and Technology (Volume 2, Issue 2) |
| DOI | 10.11648/j.ajmst.20260202.13 |
| Page(s) | 32-56 |
| Creative Commons |
This is an Open Access article, distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution and reproduction in any medium or format, provided the original work is properly cited. |
| Copyright |
Copyright © The Author(s), 2026. Published by Science Publishing Group |
Acute Respiratory Distress Syndrome, SARS - CoV -2, Ganoderma Lucidum, Alkhadaya
Autoantibodies IgG | With brain damage | No brain damage | Intact | p | Mann-Winney U- test |
|---|---|---|---|---|---|
IgG to NF 200 | 0.390 (0.283-0.653 | 0.296 (0.257-0.392) | 0.202 (0.164-0.259) | < 0.05 | 0.7633212 |
IgG to glial fibrillary acidic protein | 0.415 (0.305-0.450) | 0.254 (0.231-0.450) | 0.249 (0.209-0.363) | < 0.05 | 0.7112906 |
IgG to S 100 B | 0.722 (0.494-1.096) | 0.373 (0.241-1.096) | 0.234 (0.184-0.303) | < 0.01 | 1,2165880 |
IgG to total myelin protein | 0.237 (0.196-0.265) | 0.162 (0.141-0.209) | 0.153 (0.118-0.177) | < 0.01 | 1,3688104 |
IgG to voltage-dependent calcium channels | 0.272 (0.222-0.344) | 0.196 (0.157-0.329) | 0.152 (0.119-0.196) | < 0.01 | 0.8741105 |
IgG to H-cholinergic receptors | 0.390 (0.269-0.637) | 0.234 (0.217-0.395) | 0.176 (0.140-0.224) | < 0.01 | 0.8215549 |
IgG to glutamate receptors | 0.308 (0.284-0.544) | 0.259 (0.224-0.374) | 0.189 (0.145-0.219) | < 0.01 | 0.9155491 |
IgG to GABA receptors | 0.321 (0.284-0.621) | 0.266 (0.232-0.373 | 0.192 (0.138-0.259) | < 0.01 | 0.7610028 |
IgG to dopamine receptors | 0.303 (0.293-0.632) | 0.295 (0.232-0.397) | 0.186 (0.153-0.250) | < 0.01 | 0.9210083 |
IgG to serotonin receptors | 0.380 (0.315-0.482) | 0.349 (0.328-0.513) | 0.244 (0.202-0.303) | < 0.01 | 0.8316382 |
IgG to opiate receptors | 3.81±0.36 | 9.9±0.36 | 1.32±0.15 | < 0.05 | 0 |
IgG to beta- endorphin | 0.316 (0.296-0.344) | 0.392 (0.367-0.421) | 0.122 (0.103-0.145) | < 0.01 | 1,4399014 |
Indicators | χ 2 (Pearson criterion) | U test (Mann-Whinney test) | H test (Kraskes-Wallis test) | W -test Shapiro- Wilk test |
|---|---|---|---|---|
Main group | 0.9185001 | 0.9008417 | 0.9341006 | 0.9120318 |
Control group | 0.8210034 | 0.8521073 | 0.8438502 | 0.8230184 |
Indicators | χ 2 (Pearson criterion) | U test (Mann-Whinney test) | H test (Kraskes-Wallis test) | W -test Shapiro- Wilk test |
Main group | 0.9185001 | 0.9008417 | 0.9341006 | 0.9120318 |
Control group | 0.8210034 | 0.8521073 | 0.8438502 | 0.8230184 |
Indicators | 95% CI | OSH | Bonferroni correction | r (Spearman correlation analysis) | Weinberg equilibrium |
|---|---|---|---|---|---|
Main group | 3.0-6.3 | 0.9572104 | 0.9130047 | 0.9310082 | 0.9013862 |
Control group | 2.4-7.9 | 0.8207383 | 0.8018369 | 0.7810284 | 0.8109375 |
Indicators | 95% CI | OSH | Bonferroni correction | r (Spearman correlation analysis) | Weinberg equilibrium |
Main group | 3.0-6.3 | 0.9572104 | 0.9130047 | 0.9310082 | 0.9013862 |
Control group | 2.4-7.9 | 0.8207383 | 0.8018369 | 0.7810284 | 0.8109375 |
Autoantibodies IgG | In 3 days | In 7 days | In 14 days | p | Mann-Winney U- test | r (Spearman correlation) |
|---|---|---|---|---|---|---|
IgG to NF 200 | 0.405 (0.383-0.653) | 0.496 (0.457-0.592) | 0.607 (0.564-0.659) | < 0.001 | 0.7633212 | 0.528 |
IgG to glial fibrillary acidic protein | 0.515 (0.505-0.550) | 0.554 (0.531-0.650) | 0.649 (0.609-0.763) | < 0.001 | 0.7112906 | 0.388 |
IgG to S 100 B | 0.822 (0.794-1.096) | 0.873 (0.841-1.096) | 0.934 (0.884-1.303) | < 0.001 | 1,2165880 | 0.623 |
IgG to total myelin protein | 0.337 (0.296-0.365) | 0.362 (0.341-0.409) | 0.453 (0.418-0.477) | < 0.001 | 1,3688104 | 0.512 |
IgG to voltage-dependent calcium channels | 0.372 (0.322-0.394) | 0.396 (0.357-0.429) | 0.452 (0.419-0.496) | < 0.001 | 0.8741105 | 0.475 |
IgG to H-cholinergic receptors | 0.490 (0.469-0.637) | 0.534 (0.517-0.595) | 0.576 (0.540-0.624) | < 0.001 | 0.8215549 | 0.396 |
IgG to glutamate receptors | 0.408 (0.384-0.544) | 0.459 (0.424-0.474) | 0.489 (0.445-0.519) | < 0.001 | 0.9155491 | 0.481 |
IgG to GABA receptors | 0.421 (0.384-0.621) | 0.466 (0.432-0.473 | 0.492 (0.438-0.559) | < 0.001 | 0.7610028 | 0.962 |
IgG to dopamine receptors | 0.403 (0.393-0.632) | 0.495 (0.432-0.597) | 0.586 (0.553-0.650) | < 0.001 | 0.9210083 | 0.685 |
IgG to serotonin receptors | 0.480 (0.415-0.582) | 0.549 (0.528-0.613) | 0.644 (0.602-0.703) | < 0.001 | 0.8316382 | 0.441 |
IgG to opiate receptors | 4.81±0.36 | 10.9±0.36 | 11.32±0.15 | < 0.05 | 0.0329715 | 0.293 |
IgG to beta- endorphin | 0.416 (0.396-0.444) | 0.492 (0.467-0.521) | 0.522 (0.503-0.545) | < 0.001 | 1,4399014 | 0.851 |
Autoantibodies IgG | In 3 days | In 7 days | In 14 days | p | Mann-Winney U- test | r (Spearman correlation) |
|---|---|---|---|---|---|---|
IgG to NF 200 | 0.505 (0.383-0.653) | 0.596 (0.457-0.592) | 0.707 (0.564-0.659) | < 0.001 | 0.7633212 | 0.561 |
IgG to glial fibrillary acidic protein | 0.585 (0.505-0.550) | 0.594 (0.531-0.650) | 0.669 (0.609-0.763) | < 0.001 | 0.7112906 | 0.402 |
IgG to S 100 B | 0.873 (0.794-1.096) | 0.931 (0.841-1.096) | 0.976 (0.884-1.303) | < 0.001 | 1,2165880 | 0.684 |
IgG to total myelin protein | 0.437 (0.296-0.465) | 0.462 (0.341-0.509) | 0.553 (0.418-0.577) | < 0.001 | 1,3688104 | 0.720 |
IgG to voltage-dependent calcium channels | 0.382 (0.322-0.394) | 0.406 (0.357-0.429) | 0.462 (0.419-0.496) | < 0.001 | 0.8741105 | 0.491 |
IgG to H-cholinergic receptors | 0.500 (0.469-0.637) | 0.544 (0.517-0.595) | 0.586 (0.540-0.624) | < 0.001 | 0.8215549 | 0.496 |
IgG to glutamate receptors | 0.418 (0.384-0.544) | 0.469 (0.424-0.474) | 0.499 (0.445-0.519) | < 0.001 | 0.9155491 | 0.902 |
IgG to GABA receptors | 0.431 (0.384-0.621) | 0.476 (0.432-0.473 | 0.502 (0.438-0.559) | < 0.001 | 0.7610028 | 0.332 |
IgG to dopamine receptors | 0.413 (0.393-0.632) | 0.505 (0.432-0.597) | 0.586 (0.553-0.650) | < 0.001 | 0.9210083 | 0.704 |
IgG to serotonin receptors | 0.490 (0.415-0.582) | 0.559 (0.528-0.613) | 0.654 (0.602-0.703) | < 0.001 | 0.8316382 | 0.671 |
IgG to opiate receptors | 4.86±0.36 | 11.09±0.36 | 11.48±0.15 | < 0.05 | 0.0329715 | 0.558 |
IgG to beta- endorphin | 0.426 (0.396-0.444) | 0.502 (0.467-0.521) | 0.522 (0.503-0.545) | < 0.001 | 1,4399014 | 0.924 |
Parameter | Cutoff | AUC |
|---|---|---|
Lymphocytes, thousands/µL | 0,89 | 0,660 |
Platelets х 10҆*9 | 253,5 | 0,538 |
NLR | 4,26 | 0,784 |
PI, % | 106,4 | 0,651 |
Creatinine, µmol/L | 100,1 | 0,652 |
CRP, mg/L | 83,7 | 0,718 |
Chitotriosidase nmol/mL/h | 150 | 0,683 |
threshold values | Odds ratio | 95% confidence interval |
|---|---|---|
Lymphocytes, thousands/µL | 3,64 | 1,739-7,756 |
Platelets х 10҆*9 | 3,317 | 1,373-7,533 |
NLR | 2,667 | 0,794-8,954 |
PI, % | 3,344 | 1,54-7,3 |
Creatinine, µmol/L | 3,527 | 1,710-7,277 |
CRP, mg/L | 2,973 | 1,077-8,208 |
Chitotriosidase nmol/mL/h | 2,942 | 1,433-6,040 |
Parameters | M±SD | 95% CI | r | χ 2 |
|---|---|---|---|---|
Amplitude of the M-response of m. Biceps femoris., mV | 1.22±0.56 | 0.99-1.47 | <0.05 | 12,357 |
M-response amplitude m. triceps brachii., mV | 1.35±0.44 | 1.01-1.95 | < 0.05 | 10.98 |
The amplitude of the M-response of the peroneus muscle longus, mV | 0.61±0.22 | 0.33-1.02 | < 0.05 | 9,847 |
Amplitude of the M-response of the gastrocnemius muscle, mV | 0.65±0.27 | 0.35-1.04 | < 0.05 | 9,851 |
Amplitude of the M-response of m. extensor carpi ulnaris , mV | 0.44±0.15 | 0.23-0.56 | < 0.05 | 8.74 |
Amplitude of the M-response of m. tibialis cranialis, mV | 0.47±0.16 | 0.21-0.63 | < 0.05 | 8.63 |
Parameters | M±SD | 95% CI | r | χ 2 |
|---|---|---|---|---|
Amplitude of the M-response of m. Biceps femoris., mV | 1.92±0.56 | 1.22-2.31 | <0.05 | 10,347 |
M-response amplitude m. triceps brachii., mV | 1.85±0.44 | 1.01-2.15 | < 0.05 | 9,154 |
The amplitude of the M-response of the peroneus muscle longus, mV | 0.92±0.22 | 0.63-1.22 | < 0.05 | 9,774 |
Amplitude of the M-response of the gastrocnemius muscle, mV | 0.65±0.27 | 0.35-1.04 | < 0.05 | 8,332 |
Amplitude of the M-response of m. extensor carpi ulnaris, mV | 0.84±0.15 | 0.23-0.96 | < 0.05 | 7,891 |
Amplitude of the M-response of m. tibialis cranialis., mV | 0.97±0.16 | 0.21-0.63 | < 0.05 | 12,033 |
Parameters | M±SD | 95% CI | r | χ 2 |
|---|---|---|---|---|
Amplitude of the M-response of m. Biceps femoris., mV | 1.67±0.56 | 1.20-2.02 | <0.05 | 16,441 |
M-response amplitude m. triceps brachii., mV | 1.78±0.44 | 1.34-1.95 | < 0.05 | 13.15 |
The amplitude of the M-response of the peroneus muscle longus, mV | 0.93±0.22 | 0.63-1.12 | < 0.05 | 8,334 |
Amplitude of the M-response of the gastrocnemius muscle, mV | 0.95±0.27 | 0.75-1.23 | < 0.05 | 9,771 |
Amplitude of the M-response of m. extensor carpi ulnaris., mV | 0.87±0.15 | 0.63-0.96 | < 0.05 | 3.57 |
Amplitude of the M-response of m. tibialis cranialis., mV | 0.91±0.16 | 0.81-1.13 | < 0.05 | 9.52 |
ENMG parameter | Indicators (after 14 days) |
|---|---|
Amplitude of the M-response of the Biceps muscle femoris (n.medianus) | 2.5±1.9 |
DL n.medianus | 4.8±3.1 |
SRV n. medianus | 23.2±9.0 |
Amplitude of the M-response with m. gastrocnemius (n.tibialis) | 1.4±0.1 |
DL n.tibialis | 6.5±3.2 |
CRV n.tibialis | 20.7±6.9 |
No M-response with m. gastrocnemius (n.tibialis) | 38.8% (n=7) |
Absence of PD n.suralis | 100% (n=18) |
EMG | Electromyography |
n. | Nerves |
| [1] | Abilov PM, Makhkamova FT (2018) Clinical and functional evaluation of the effectiveness of treatment of chronic catarrhal gingivitis in children with the use of biologically active additives based on Ganoderma Lutsidum. Pediatrics, Scientific and Practical Journal, Tashkent 1: 108-111. |
| [2] | Abilov PM Efficienty of individual prophylaxis of Dental Caries using dental gel Ispring based on Ganoderma Lucidum in schoolchildren in Tashkent. Journal of Dentistry and Oral Health. 2018; 5: 1 -4. |
| [3] | Abilov PM, Iriskulov BU, Saydalikhodjaeva OZ, Seytkarimova GS, Norboeva SA, Musaev Kh. A. An experimental assessment of the influence of Ganoderma lucidum on the state of oxidative stress. International journal of scientific & technology research 2020; 9(3): 6645-6649. |
| [4] | Abdullah, F., Myers, J., Basu, D., Tintinger, G., Ueckermann, V., Mathebula, M., et al. (2022). Decreased severity of disease during the first global omicron variant covid-19 outbreak in a large hospital in tshwane, South Africa. Int. J. Infect. Dis. 116, 38-42. |
| [5] | Abu- Raddad, L. J., Chemaitelly, H., and Butt, A. A. National Study Group for COVID-19 Vaccination (2021). Effectiveness of the BNT162b2 Covid-19 vaccine against the B. 1.1.7 and B. 1.351 variants. N. Engl. J. Med. 385, 187-189. |
| [6] | Ackermann, M., Verleden, SE, Kuehnel, M., Haverich, A., Welte, T., Laenger, F., et al. (2020). Pulmonary vascular Endothelialitis, thrombosis, and angiogenesis in Covid-19. N. Engl. J. Med. 383, 120-128. |
| [7] | Afkhami, S., D'Agostino, M. R., Zhang, A., Stacey, H. D., Marzok, A., Kang, A., et al. (2022). Respiratory mucosal delivery of next-generation COVID-19 vaccine provides robust protection against both ancestral and variant strains of SARS-CoV-2. Cell 185, 896-915. |
| [8] | Alonso- Canovas, A., Kurtis, M. M., Gomez- Mayordomo, V., Macías-García, D., Gutiérrez Viedma, A., Mondragón Rezola, E., et al. (2023). Functional neurological disorders after COVID-19 and SARS-CoV-2 vaccines: a national multicentre observational study. J. Neurol. Neurosurg. Psychiatry 94, 776-777. |
| [9] | Alu, A., Chen, L., Lei, H., Wei, Y., Tian, X., and Wei, X. (2022). Intranasal COVID-19 vaccines: from bench to bed. EBioMedicine 76: 103841. |
| [10] | Andreano, E., Piccini, G., Licastro, D., Casalino, L., Johnson, N. V., Paciello, I., et al. (2021). SARS-CoV-2 escape from a highly neutralizing COVID-19 convalescent plasma. Proc. Natl. Acad. Sci. USA 118: 54118. |
| [11] | Ao, D., He, X., Liu, J., and Xu, L. (2023). Strategies for the development and approval of COVID-19 vaccines and therapeutics in the post-pandemic period. Signal Transduct. Target. Ther. 8: 466. |
| [12] | Arvin, A. M., Fink, K., Schmid, M. A., Cathcart, A., Spreafico, R., Havenar-Daughton, C., et al. (2020). A perspective on potential antibody-dependent enhancement of SARS-CoV-2. Nature 584, 353-363. |
| [13] | Baum, A., Fulton, B. O., Wloga, E., Copin, R., Pascal, K. E., Russo, V., et al. (2020). Antibody cocktail to SARS-CoV-2 spike protein prevents rapid mutational escape seen with individual antibodies. Science 369, 1014-1018. |
| [14] | Berardicurti, O., Ruscitti, P., Ursini, F., D'Andrea, S., Ciaffi, J., Meliconi, R., et al. (2020). Mortality in tocilizumab -treated patients with COVID-19: a systematic review and meta-analysis. Clin. Exp. Rheumatol. 38, 1247-1254. |
| [15] | Blanco- Melo, D., Nilsson- Payant, B. E., Liu, W. C., Uhl, S., Hoagland, D., Møller, R., et al. (2020). Imbalanced host response to SARS-CoV-2 drives development of COVID-19. Cell 181, 1036-1045. |
| [16] | Boehm, E., Kronig, I., Neher, R. A., Eckerle, I., Vetter, P., Kaiser, L., et al. (2021). Novel SARS-CoV-2 variants: the pandemics within the pandemic. Clin. Microbiol. Infect. 27, 1109-1117. |
| [17] | Borczuk, A. C. (2021). Pulmonary pathology of COVID-19: a review of autopsy studies. Curr. Opin. Pulm. Med. 27, 184-192. |
| [18] | Bourgonje, AR, Abdulle, AE, Timens, W., Hillebrands, JL, Navis, GJ, Gordijn, S. J., et al. (2020). Angiotensin-converting enzyme 2 (ACE2), SARS-CoV-2 and the pathophysiology of coronavirus disease 2019 (COVID-19). J. Pathol. 251, 228-248. |
| [19] | Bournazos, S., Gupta, A., and Ravetch, J. V. (2020). The role of IgG fc receptors in antibody-dependent enhancement. Nat. Rev. Immunol. 20, 633-643. |
| [20] | Bricker, T. L., Darling, T. L., Hassan, A. O., Harastani, H. H., Soung, A., Jiang, X., et al. (2020). A single intranasal or intramuscular immunization with chimpanzee adenovirus vectored SARS-CoV-2 vaccine protects against pneumonia in Hamsters. bioRxiv [Preprint]. |
| [21] | Bukhari K, Mulley G, Gulyaeva AA, et al. Description and initial characterization of metatranscriptomic nidovirus -like genomes from the proposed new family abyssoviridae, and from a sister group to the coronavirinae, the proposed genus alphaletovirus. Virology. 2018; 524: 160-171. |
| [22] | Butowt, R., Bilinska, K., and Von Bartheld, C. S. (2020). Chemosensory dysfunction in COVID-19: integration of genetic and epidemiological data points to D614G spike protein variant as a contributing factor. ACS Chem. Neurosci. 11, 3180-3184. |
| [23] | Cameroni, E., Bowen, J. E., Rosen, L. E., Saliba, C., Zepeda, S. K., Culap, K., et al. (2022). Broadly neutralizing overcome antibodies SARS-CoV-2 omicron antigenic shift. Nature 602, 664-670. |
| [24] | Campos, K. R., Sacchi, C. T., Abbud, A., and Caterino -de-Araujo, A. (2021). SARSCoV-2 variants in severely symptomatic and deceased persons who had been vaccinated against COVID-19 in São Paulo, Brazil. Rev. Panam. Salud Publica 45: e 126. |
| [25] | Cao, Z., Gao, W., Bao, H., Feng, H., Mei, S., Chen, P., et al. (2023). VV116 versus Nirmatrelvir -ritonavir for Oral treatment of Covid-19. N. Engl. J. Med. 388, 406-417. |
| [26] | Carvajal JJ, Garc´ıa -Castillo V, Cuellar SV, Campillay-Ve´liz CP, Salazar -Ardiles C, Avellaneda AM, Muñoz CA, Retamal-D´ıaz A, Bueno S. M, Gonza´lez PA, Kalergis AM and Lay MK (2024) New insights into the pathogenesis of SARS-CoV- 2 during and after the COVID-19 pandemic. Front. Immunol. 15: 1363572. |
| [27] | Chemaitelly, H., Yassine, H. M., Benslimane, F. M., al Khatib, H. A., Tang, P., Hasan, M. R., et al. (2021). mRNA-1273 COVID-19 vaccine effectiveness against the B. 1.1.7 and B. 1.351 variants and severe COVID-19 disease in Qatar. Nat. Med. 27, 1614-1621. |
| [28] | Chen, J., Wang, P., Yuan, L., Zhang, L., Zhang, L., Zhao, H., et al. (2022). A live attenuated virus-based intranasal COVID-19 vaccine provides rapid, prolonged, and broad protection against SARS-CoV-2. Sci. Bull. 67, 1372-1387. |
| [29] | Choudhary, O. P., Priyanka, M. T. A., Mohammed, T. A., and Singh, I. (2021). Intranasal COVID-19 vaccines: is it a boon or bane? Int. J. Surg. 94: 106119. |
| [30] | Chu, H., Chan, J. F., Wang, Y., Yuen, T. T., Chai, Y., Hou, Y., et al. (2020). Comparative replication and immune activation profiles of SARS-CoV-2 and SARS- CoV in human lungs: an ex vivo study with implications for the pathogenesis of COVID-19. Clin. Infect. Dis. 71, 1400-1409. |
| [31] | Colson, P., Bader, W., Fantini, J., Dudouet, P., Levasseur, A., Pontarotti, P., et al. (2023). From viral democratic genomes to viral wild bunch of quasispecies. J. Med. Virol. 95: e 29209. |
| [32] | Costa, R., Olea, B., Bracho, M. A., Albert, E., de Michelena, P., Martínez -Costa, C., et al. (2022). RNA viral loads of SARS-CoV-2 alpha and Delta variants in nasopharyngeal specimens at diagnosis stratified by age, clinical presentation and vaccination status. J. Infect. 84, 579-613. |
| [33] | Davies, N. G., Abbott, S., Barnard, R. C., Jarvis, C. I., Kucharski, A. J., Munday, J. D., et al. (2021). Estimated transmissibility and impact of SARS-CoV-2 lineage B. 1.1.7 in England. Science 372: 3055. |
| [34] | de Melo, G. D., Lazarini, F., Levallois, S., Hautefort, C., Michel, V., Larrous, F., et al. (2021). COVID-19-related anosmia is associated with viral persistence and inflammation in human olfactory epithelium and brain infection in Hamsters. Sci. Transl. Med. 13: 8396. |
| [35] | de Wit, E., van Doremalen, N., Falzarano, D., and Munster, V. J. (2016). SARS and MERS: recent insights into emerging coronaviruses. Nat. Rev. Microbiol. 14, 523-534. |
| [36] | Dejnirattisai, W., Huo, J., Zhou, D., Zahradník, J., Supasa, P., Liu, C., et al. (2022). SARS-CoV-2 omicron -B. 1.1.529 leads to widespread escape from neutralizing antibody responses. Cell 185, 467-484. |
| [37] | Delgado, S., Somovilla, P., Ferrer -Orta, C., Martínez -González, B., Vázquez- Monteagudo, S., Muñoz-Flores, J., et al. (2024). Incipient functional SARSCoV-2 diversification identified through neural network haplotype maps. Proc. Natl. Acad. Sci. USA 121: e 2317851121. |
| [38] | Domizio, J. D., Gulen, M. F., Saidoune, F., Thacker, V. V., Yatim, A., Sharma, K., et al. (2022). The cGAS -STING pathway drives type I IFN immunopathology in COVID-19. Nature 603, 145-151. |
| [39] | Du, L., He, Y., Zhou, Y., Liu, S., Zheng, B. J., and Jiang, S. (2009). The spike protein of SARS-CoV-2 is a target for vaccine and therapeutic development. Nat. Rev. Microbiol. 7, 226-236. |
| [40] | Ellul, M. A., Benjamin, L., Singh, B., Lant, S., Michael, B. D., Easton, A., et al. (2020). Neurological associations of COVID-19. Lancet Neurol. 19, 767-783. |
| [41] | Fan, Y., Li, X., Zhang, L., Wan, S., Zhang, L., and Zhou, F. (2022). SARS-CoV-2 omicron variant: recent progress and future perspectives. Signal Transduct. Target. Ther. 7: 141. |
| [42] | Faria, NR, Mellan, TA, Whittaker, C., Claro, IM, Candido, DDS, Mishra, S., et al. (2021). Genomics and epidemiology of the P. 1 SARS-CoV -2 lineage in Manaus, Brazil. Science 372, 815-821. |
| [43] | Fernández-Castañeda, A., Lu, P., Geraghty, A. C., Song, E., Lee, M. H., Wood, J., et al. (2022). Mild respiratory COVID can cause multi-lineage neural cell and myelin dysregulation. Cell 185, 2452-2468. |
| [44] | Fiolet, T., Kherabi, Y., MacDonald, C. J., Ghosn, J., and Peiffer-Smadja, N. (2022). Comparing COVID-19 vaccines for their characteristics, efficacy and effectiveness against SARS-CoV-2 and variants of concern: a narrative review. Clin. Microbiol. Infect. 28, 202-221. |
| [45] | Fisman, D. N., and Tuite, A. R. (2021). Evaluation of the relative virulence of novel SARS-CoV-2 variants: a retrospective cohort study in Ontario, Canada. J Assoc Med Can 193, E1619-E1625. |
| [46] | Forchette, L., Sebastian, W., and Liu, T. (2021). A comprehensive review of COVID-19 virology, vaccines, variants, and therapeutics. Curr Med Sci. 41, 1037-1051. |
| [47] | Gao, Y. D., Ding, M., Dong, X., Zhang, J. J., Kursat Azkur, A., Azkur, D., et al. (2021). Risk factors for severe and critically ill COVID-19 patients: a review. Allergy 76, 428-455. |
| [48] | Garcia-Beltran, W. F., Lam, E. C., St. Denis, K., Nitido, A. D., Garcia, Z. H., Hauser, B. M., et al. (2021). Multiple SARS-CoV-2 variants escape neutralization by vaccine-induced humoral immunity. Cell 184, 2372-2383. |
| [49] | Georg, P., Astaburuaga-García, R., Bonaguro, L., Brumhard, S., Michalick, L., Lippert, L. J., et al. (2022). Complement activation induces excessive T cell cytotoxicity in severe COVID-19. Cell 185, 493-512. |
| [50] | GeurtsvanKessel, CH, Geers, D., Schmitz, KS, Mykytyn, AZ, Lamers, MM, Bogers, S., et al. (2022). Divergent SARS-CoV-2 omicron-reactive T and B cell responses in COVID-19 vaccine recipients. Sci Immunol. 7: 2202. |
| [51] | Greaney, AJ, Loes, AN, Crawford, KHD, Starr, TN, Malone, KD, Chu, HY, et al. (2021). Comprehensive mapping of mutations in the SARS-CoV-2 receptorbinding domain that affect recognition by polyclonal human plasma antibodies. Cell Host Microbe 29, 463-476. |
| [52] | Greene, C., Connolly, R., Brennan, D., Laffan, A., O'Keeffe, E., Zaporojan, L., et al. (2024). Blood-brain barrier disruption and sustained systemic inflammation in individuals with long COVID-associated cognitive impairment. Nat. Neurosci. 27, 421-432. |
| [53] | Gruell, H., Vanshylla, K., Tober -Lau, P., Hillus, D., Schommers, P., Lehmann, C., et al. (2022). mRNA booster immunization elicits potent neutrali zing serum activity against the SARS-CoV-2 omicron variant. Nat. Med. 28, 477-480. |
| [54] | Gupta, A., Madhavan, M. V., Sehgal, K., Nair, N., Mahajan, S., Sehrawat, T. S., et al. (2020). Extrapulmonary manifestations of COVID-19. Nat. Med. 26, 1017-1032. |
| [55] | Hadj, H. I. (2022). Covid-19 vaccines and variants of concern: a review. Rev. Med. Virol. 32: e 2313. |
| [56] | Hadj Hassine, I., Ben M'hadheb, M., and Menéndez-Arias, L. (2022). Lethal mutagenesis of RNA viruses and approved drugs wit h antiviral mutagenic activity. Viruses 14: 841. |
| [57] | Hadjadj, J., Yatim, N., Barnabei, L., Corneau, A., Boussier, J., Smith, N., et al. (2020). Impaired type I interferon activity and inflammatory responses in severe COVID-19 patients. Science 369, 718-724. |
| [58] | Harvey, W. T., Carabelli, A. M., Jackson, B., Gupta, R. K., Thomson, E. C. Harrison, E. M., et al. (2021). SARS-CoV-2 variants, spike mutations and immune escape. Nat. Rev. Microbiol. 19, 409-424. |
| [59] | Hashimoto, T., Perlot, T., Rehman, A., Trichereau, J., Ishiguro, H., Paolino, M., et al. (2012). ACE2 links amino acid malnutrition to microbial ecology and intestinal inflammation. Nature 487, 477-481. |
| [60] | Hassan, AO, Kafai, NM, Dmitriev, IP, Fox, JM, Smith, BK, Harvey, IB, et al. (2020). A single-dose intranasal ChAd vaccine protects upper and lower respiratory tracts against SARS-CoV-2. Cell 183, 169-184. |
| [61] | Heurich, A., Hofmann-Winkler, H., Gierer, S., Liepold, T., Jahn, O., and Pöhlmann, S. (2014). TMPRSS2 and ADAM17 cleave ACE2 differentially and only proteolysis by TMPRSS2 augments entry driven by the severe acute respiratory syndrome coronavirus spike protein. J. Virol. 88, 1293-1307. |
| [62] | Ho, C. Y., Salimian, M., Hegert, J., O'Brien, J., Choi, S. G., Ames, H., et al. (2022). Postmortem assessment of olfactory tissue degeneration and microvasculopathy in patients with COVID-19. JAMA Neurol. 79, 544-553. |
| [63] | Hoffmann, M., Kleine -Weber, H., Schroeder, S., Krüger, N., Herrler, T., Erichsen, S., et al. (2020). SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 181, 271-280. |
| [64] | Holter, J. C., Pischke, S. E., de Boer, E., Lind, A., Jenum, S., Holten, A. R., et al. (2020). Systemic complement activation is associated with respiratory failure in COVID-19 hospitalized patients. Proc. Natl. Acad. Sci. USA 117, 25018-25025. |
| [65] | Huang, C., Wang, Y., Li, X., Ren, L., Zhao, J., Hu, Y., et al. (2020). Clinical features of patients infected with 2019 novel coronavirus in Wuhan. Lancet 395, 497-506. |
| [66] | Hue, S., Beldi-Ferchiou, A., Bendib, I., Surenaud, M., Fourati, S., Frapard, T., et al. (2020). Uncontrolled innate and impaired adaptive immune responses in patients with COVID-19 acute respiratory distress syndrome. Am. J. Respira. Crit. Care Med. 202, 1509-1519. |
| [67] | Hui, KPY, Ho, JCW, Cheung, MC, Ng, KC, Ching, RHH, Lai, KL, et al. (2022). SARS-CoV-2 omicron variant replication in human bronchus and lung ex vivo. Nature 603, 715-720. |
| [68] | Jingshu Zhang, Melanie Rissmann, Thijs Kuiken, and Bart L. Haagmans Comparative Pathogenesis of Severe Acute Respiratory Syndrome Coronaviruses. Annu. Rev. Pathol. Mech. Dis. 2024. 19: 423-51. |
| [69] | Li X, Mi Z, Liu Z and Rong P (2024) SARS-CoV-2: pathogenesis, therapeutics, variants, and vaccines. Front. Microbiol. 15: 1334152. |
| [70] | Natalie M. Kirk, Yuying Liang & Hinh Ly (2024) Pathogenesis and virulence of coronavirus disease: Comparative pathology of animal models for COVID-19, Virulence, 15: 1, 2316438, |
| [71] | Zhang, X., Wu, S., Wu, B., Yang, Q., Chen, A., Li, Y., et al. (2021). SARS-CoV-2 omicron strain exhibits potent capabilities for immune evasion and viral entrance. Signal Transduct. Target. Ther. 6: 430. |
APA Style
Mеlisovich, A. P. (2026). A Comprehensive Assessment of Post-COVID-19 Changes and Its Correction with the Help of G. Lucidum and Alkhaday. American Journal of Medical Science and Technology, 2(2), 32-56. https://doi.org/10.11648/j.ajmst.20260202.13
ACS Style
Mеlisovich, A. P. A Comprehensive Assessment of Post-COVID-19 Changes and Its Correction with the Help of G. Lucidum and Alkhaday. Am. J. Med. Sci. Technol. 2026, 2(2), 32-56. doi: 10.11648/j.ajmst.20260202.13
@article{10.11648/j.ajmst.20260202.13,
author = {Abilov Pulat Mеlisovich},
title = {A Comprehensive Assessment of Post-COVID-19 Changes and Its Correction with the Help of G. Lucidum and Alkhaday},
journal = {American Journal of Medical Science and Technology},
volume = {2},
number = {2},
pages = {32-56},
doi = {10.11648/j.ajmst.20260202.13},
url = {https://doi.org/10.11648/j.ajmst.20260202.13},
eprint = {https://article.sciencepublishinggroup.com/pdf/10.11648.j.ajmst.20260202.13},
abstract = {In December 2019, a series of acute respiratory illnesses characterized by rapid worsening of symptoms, pneumonia, acute respiratory distress syndrome (ARDS), and in some cases, death were observed in Wuhan City, Hubei Province, China. To achieve the set objective, the results of treatment of 100 sexually mature rats of both sexes with coronavirus infection caused by COVID -19 were analyzed. All animals were divided into groups: Group 1 - animals with coronavirus infection with a confirmed positive PCR test, treated with ivermectin at a dosage of 300 mg of body weight (n = 25), Group 2 - animals with coronavirus infection treated with baicalin at a dosage of 500 mg (n = 25), Group 3 - animals with coronavirus infection treated with molnupiravir 25 mg / kg of body weight (n = 25), Group 4 - animals with coronavirus infection treated with a new drug based on G. lucidum and black cumin (n = 25). Lymphocytic myocarditis was detected in all rats (100%). Dystrophic changes in cardiomyocytes were also detected in 57.4% of cases, apoptotic bodies in 29.41% of cases, uneven cardiomyocyte hypertrophy in 85.29% of cases, and lipofuscinosis in 45.59% of cases. Subendocardial lipomatosis in the cardiac conduction system was observed in 51.47% of cases. The average percentage of lipomatosis area from the micrograph area was 2.47±3.12%. The area under the curve (AUC) = 0.91 (95% CI = 0.85-0.98). The prediction accuracy of the prognostic model was 91.7%, sensitivity was 0.87%, and specificity was 0.95%. AUC was 0.91 (95% CI = 0.85-0.98), p < 0.001.},
year = {2026}
}
TY - JOUR T1 - A Comprehensive Assessment of Post-COVID-19 Changes and Its Correction with the Help of G. Lucidum and Alkhaday AU - Abilov Pulat Mеlisovich Y1 - 2026/03/10 PY - 2026 N1 - https://doi.org/10.11648/j.ajmst.20260202.13 DO - 10.11648/j.ajmst.20260202.13 T2 - American Journal of Medical Science and Technology JF - American Journal of Medical Science and Technology JO - American Journal of Medical Science and Technology SP - 32 EP - 56 PB - Science Publishing Group UR - https://doi.org/10.11648/j.ajmst.20260202.13 AB - In December 2019, a series of acute respiratory illnesses characterized by rapid worsening of symptoms, pneumonia, acute respiratory distress syndrome (ARDS), and in some cases, death were observed in Wuhan City, Hubei Province, China. To achieve the set objective, the results of treatment of 100 sexually mature rats of both sexes with coronavirus infection caused by COVID -19 were analyzed. All animals were divided into groups: Group 1 - animals with coronavirus infection with a confirmed positive PCR test, treated with ivermectin at a dosage of 300 mg of body weight (n = 25), Group 2 - animals with coronavirus infection treated with baicalin at a dosage of 500 mg (n = 25), Group 3 - animals with coronavirus infection treated with molnupiravir 25 mg / kg of body weight (n = 25), Group 4 - animals with coronavirus infection treated with a new drug based on G. lucidum and black cumin (n = 25). Lymphocytic myocarditis was detected in all rats (100%). Dystrophic changes in cardiomyocytes were also detected in 57.4% of cases, apoptotic bodies in 29.41% of cases, uneven cardiomyocyte hypertrophy in 85.29% of cases, and lipofuscinosis in 45.59% of cases. Subendocardial lipomatosis in the cardiac conduction system was observed in 51.47% of cases. The average percentage of lipomatosis area from the micrograph area was 2.47±3.12%. The area under the curve (AUC) = 0.91 (95% CI = 0.85-0.98). The prediction accuracy of the prognostic model was 91.7%, sensitivity was 0.87%, and specificity was 0.95%. AUC was 0.91 (95% CI = 0.85-0.98), p < 0.001. VL - 2 IS - 2 ER -
Department of Normal and Pathological Physiology, Tashkent State Medical University, Tashkent, Uzbekistan

Figure 1. Changes in the myocardium of rats under the influence of coronavirus infection.

Figure 2. Endocardial sclerosis detected in rats exposed to coronavirus infection.

Figure 3. Immunohistochemical reactions for detection of CD45 in animals with coronavirus infection.

Figure 4. CD3+ lymphocytes in rats with coronavirus infection.
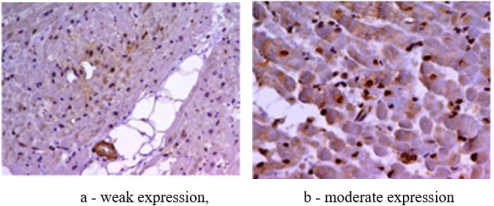
Figure 5. TLR type 4 in experimental animals with coronavirus infection.
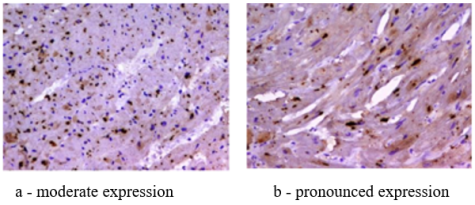
Figure 6. TLR type 9 in experimental animals with coronavirus infection.

Figure 7. Expression of the SARS-CoV-2 spike protein.

Figure 8. Expression of SARS-CoV-2 nucleocapsid protein.
Figure 9. Application of multivariate ANOVA analysis in coronavirus infection with and without spinal cord injury.

Figure 10. The ratio of IgG to the severity of the inflammatory process in coronavirus infection with and without brain damage.

Figure 11. ROC analysis of the possible influence of the ELIN test on the course of coronavirus infection.

Figure 12. Kaplan-Meier curve in groups: 1-main group, 0-control group.

Figure 13. Correlation matrix autoantibody titer IgG to various antigens and receptors (ELIN test) in the dynamics of observation in animals of the main group.

Figure 14. Correlation matrix autoantibody titer IgG to various antigens and receptors (ELIN test) in the dynamics of observation in animals of the control group.

Figure 15. Nonparametric indicators of the ELIN test for coronavirus infection based on the Kraskes-Wallis criterion with Bonferroni correction.

Figure 16. One-way regression analysis of the influence of IgG to various antigens and receptors (ELIN test) in the dynamics of observation in animals of the main group.

Figure 17. One-way regression analysis of the influence of IgG to various antigens and receptors (ELIN test) in the dynamics of observation in animals of the control group.
Figure 18. Determination of the influence of IgG to various antigens and receptors (ELIN test) in the dynamics of observation using the long-rank test.

Figure 19. Correlation of one-way regression analysis and long - rang test on the experimental model of coronavirus infection and evaluation the influence of IgG to various antigens and receptors (ELIN test) in the dynamics of observation.

Figure 20. Survival analysis depending on the initial chitotriosidase activity.
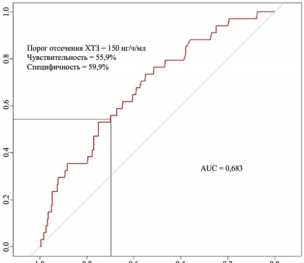
Figure 21. ROC curve characterizing the dependence of the probability of developing a fatal outcome in surviving rats with COVID-19 on the value of the logistic function P.
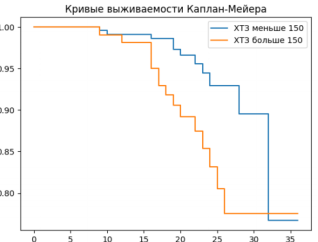
Figure 22. Kaplan -Meier curves showing the survival of rats with coronavirus infection with chitotriosidase activity above and below 150 nmol /ml/h.

